Мобилизация жира
В настоящее время, после открытия в 2004 году адипоцитарной триглицеридлипазы (ATGL, adipose triglyceride lipase, деснутрин), прежняя концепция липолиза получила новое прочтение. ATGL несет ответственность за большую часть липолитической активности как в базальных, так и в стимулированных условиях.
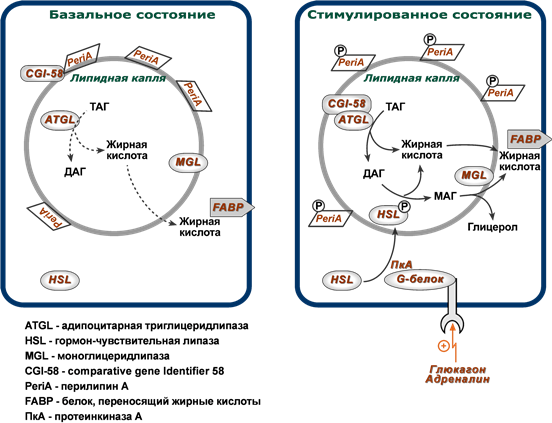
В обычных условиях на поверхности липидной капли находятся белок перилипин, ATGL и ее коактиватор под названием CGI-58 (comparative gene identification-58, или также Abhd5, abhydrolase domain-containing 5).
Гормон-чувствительная липаза (HSL, hormone sensitive lipase) и белок, переносящий жирные кислоты (БПЖК, FABP, fatty acid binding protein) в это время преимущественно находятся в цитоплазме.
В спокойном адипоците
- ATGL отщепляет жирные кислоты (С16-С18) во 2-м положении триацилглицерола с образованием свободной жирной кислоты и 1,3-ДАГ. Происходит базальный липолиз.
В стимулированном адипоците
- перилипин, фосфорилируемый протеинкиназой А, изменяет свою конформацию и уходит от поверхности липидной капли в цитозоль.
- белок CGI-58 отделяется от перилипина и присоединяется к ATGL, формируя с ней активный комплекс и меняя ее специфичность.
- триглицеридлипаза (ATGL), после связывания с CGI-58, начинает отщеплять жирные кислоты от 1-го положения триацилглицеролов, продуцируя 2,3-ДАГ. Именно 2,3-ДАГ является субстратом гормон-чувствительной липазы (HSL).
- гормон-чувствительная липаза (HSL) фосфорилируется протеинкиназой А и/или протеинкиназой G и в такой активной форме мигрирует к липидной капле. Отсутствие перилипина позволяет этой липазе подобраться к ядру липидной капли, где она гидролизует 2,3-диацилглицеролы до 2-моноацилглицеролов и свободной жирной кислоты.
- МАГ-липаза находится как в цитозоле, так и на поверхности липидной капли и гидролизует моноацилглицеролы.
Хотя главным субстратом HSL являются диацилглицеролы, также она может гидролизовать ТАГ и МАГ, в соотношении скоростей гидролиза 20 : 2 : 1 для ДАГ:ТАГ:МАГ соответственно.
Жирные кислоты покидают жировую клетку и, связываясь с сывороточным альбумином, переносятся кровотоком. Затем они освобождаются от альбумина и с помощью тканевых белков, переносящих жирные кислоты, проникают в мишеневые клетки.
Регуляция липолиза в адипоците
Субстратный цикл
Базальный распад и синтез триацилглицеролов в жировой ткани объединены в субстратный цикл, который заключается в том, что 1,3-ДАГ, продукт малоактивной ATGL, является предпочтительным субстратом одного из ферментов синтеза триацилглицеролов – ДАГ-ацил-трансферазы. В результате ATGL и ДАГ-ацил-ТФ действуют скоординированно и поддерживают непрерывный цикл гидролиза-реэтерификации ТАГ.
В состоянии покоя около 70% жирных кислот, высвобожденных при участии малоактивной ATGL, не покидают клетку и включаются обратно в состав ТАГ. Остальные 30% жирных кислот выходят в кровь. Вышедшие в кровь жирные кислоты используются в тканях, и их часть неминуемо попадает в печень, где они реэтерифицируются в ТАГ, далее формируется их транспортная форма ЛПОНП. При метаболизме последних в плазме крови образуются остаточные ЛПОНП, некоторая часть которых превращается в синусоидах печени в атерогенные ЛПНП. Нетрудно представить, что количество образуемых ЛПОНП и ЛПНП напрямую зависит от жировой массы, что связывает ожирение и развитие атеросклероза.
Активации субстратного цикла жирных кислот также способствует часто наблюдающийся дисбаланс между потреблением насыщенных и полиненасыщенных жирных кислот, так как липидная капля в адипоците окружена монослоем фосфолипидов, которые должны содержать ПНЖК. При нарушении синтеза фосфолипидов доступ ATGL-липазы к триацилглицеролам облегчается и их гидролиз ускоряется, возрастает доля насыщенных ЖК, выходящих в кровь.
Избыток насыщенных ЖК крови легко переходит в мембраны клеток эндотелия сосудов и гладких миоцитов и задерживается здесь, образуя кластеры. Это меняет свойства мембраны, нарушает ионные потоки, что повышает тонус гладких мышц, ухудшает взаимодействие рецепторов с их лигандами.