Медленная компенсация сдвигов величины pH обеспечивается физиологическими системами - регулируемо через легкие и почки, пассивно при участии костной ткани и печени.
Удаление кислоты через легкие
Газообмен в легких заключается в постоянном обмене кислородом и углекислым газом между двумя соприкасающимися компартментами – легочными альвеолами и кровеносными капиллярами. (см также Обмен кислорода и углекислого газа в легких)
Углекислый газ быстро диффундирует из плазмы и эритроцитов в альвеолярное пространство. В то же время в альвеолярном воздухе имеется высокая концентрация кислорода, который, проникая в эритроциты, образует оксигемоглобин – более сильную кислоту, чем угольная. "Кислый" оксигемоглобин отдает ионы Н+ в ассоциацию с ионом HCO3–, образуется H2CO3, которая вступает в карбоангидразную реакцию с появлением СО2, удаляемого в плазму и далее в альвеолы.
Легочная вентиляция обеспечивает удаление CO2 (или, что то же самое H2CO3) и повышение уровня pH. Она эффективна только в тесном контакте с гемоглобиновой и бикарбонатной буферной системами.
Первым фактором для активации дыхательной системы является концентрация ионов Н+, влияющих на периферические (каротидные тельца) и центральные (продолговатый мозг) рецепторы. Закисление крови уже через 1‑2 минуты вызывает стимуляцию дыхательного центра, повышая его активность в 4‑5 раз. И наоборот, снижение кислотности крови понижает активность дыхательного центра в 2‑4 раза.
Вторым фактором регуляции служит концентрация CO2. Чувствительные хеморецепторы для CO2 находятся в продолговатом мозге, в аортальном и каротидных тельцах. В случае хронической гиперкапнии (увеличении CO2) рецепторы дыхательного центра теряют чувствительность к CO2 (десенсибилизация). При этом основным стимулом становится pO2.
Третий регулятор вентиляции – сдвиги концентрации O2. На pO2 реагируют, главным образом, периферические рецепторы каротидного синуса и дуги аорты. Они являются значимыми для легочной вентиляции только при сильном снижении рО2 в артериальной крови (60-70% от нормы).
Удаление кислоты через почки
Почки играют главную роль в регуляции уровня рН плазмы крови при длительной компенсации. Движущей силой в почечных влияниях на уровень pH являются потоки ионов Na+. Ионы Na+ стремятся внутрь клеток по концентрационному градиенту, который создается при работе фермента Na+/К+‑АТФазы на базолатеральной мембране. Движение ионов Na+ из просвета канальца внутрь эпителиоцита приводит к выходу через Na+/H+‑обменник апикальной мембраны в просвет канальца ионов H+.
Благодаря почкам регулируется концентрация ионов HCO3‑ в плазме крови и ионы Н+ удаляются в мочу. Благодаря работе почек рН мочи в состоянии снижаться до 4,5-5,2. Здесь активно протекают три процесса, связанных с уборкой кислых эквивалентов:
- Реабсорбция бикарбонатных ионов HCO3-.
- Ацидогенез – удаление ионов Н+ с титруемыми кислотами (в основном в составе дигидрофосфатов NaH2PO4).
- Аммониегенез – удаление ионов Н+ в составе ионов аммония NH4+.
Реабсорбция бикарбонат-ионов
Ионы HCO3–, находящиеся в плазме крови, непрерывно фильтруются и оказываются в первичной моче. Одновременно в канальцевую жидкость при участии Na+/H+‑обменника из эпителиоцитов активно выводятся ионы H+ в обмен на ионы Na+.
Секретируемые ионы Н+ и ионы HCO3–, находящиеся в первичной моче, ассоциируют до угольной кислоты Н2СО3, которая в гликокаликсе при участии карбоангидразы распадается на СО2 и H2O. Используя возникающий градиент концентрации между просветом канальцев и цитозолем эпителиоцитов, СО2 диффундирует в клетки. Внутриклеточная карбоангидраза направляет входящий СО2 для образования угольной кислоты. После диссоциации кислоты ионы НСО3– выносятся в кровь, подщелачивая ее, ионы Н+ секретируются обратно в мочу. В итоге происходит обратное всасывание ионов НСО3–, при этом каждый добавляемый в плазму НСО3– соответствует экскреции одного иона Н+.
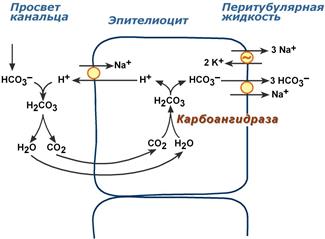
Реабсорбция бикарбонат-ионов
Пороговое значение реабсорбции бикарбонатов соответствует их нормальной концентрации в крови. Это значит, что при превышении нормального уровня в крови бикарбонаты не успеют реабсорбироваться и останутся в моче.
В проксимальных канальцах реабсорбируется 80% профильтрованного НСО3–. В целом в почечных канальцах происходит реабсорбция более 99% от фильтруемого бикарбонат-иона.
Ацидогенез
Ацидогенез заключается во взаимодействии ионов H+, секретируемых из эпителиоцитов, с анионами HРO42–. В результате в первичной моче образуется дигидрофосфат H2РO4–.
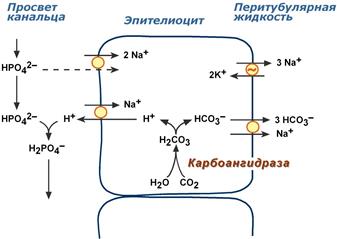
Реакции ацидогенеза
В процессе ацидогенеза в сутки выделяется 10-30 ммоль кислотных веществ, названных титруемыми кислотами. Фосфаты, являясь одной из этих кислот, играют роль буферной системы в моче. Роль ее состоит в экскреции кислых эквивалентов без потерь бикарбонат-ионов за счет дополнительного иона H+ в составе выводимого H2РO4– (по сравнению с HCO3):
HРO42– + Н2СО3 → H2РO4– + НСО3
Бикарбонат натрия в почечных канальцах реабсорбируется, а кислотно-основная реакция мочи зависит только от содержания дигидрофосфата. Хотя в крови соотношение HРO42–/ H2РO4– равно 4:1, начиная от клубочкового фильтрата к дистальным канальцам оно может достигать 50-кратного перевеса доли ионов H2РO4– (1:50).
Аммониегенез
Глутамин и глутаминовая кислота, попадая в клетки канальцев, быстро дезаминируются глутаминазой и глутаматдегидрогеназой с образованием аммиака. Являясь гидрофобным соединением, аммиак диффундирует в просвет канальца и связывает экскретируемые ионы Н+ с образованием иона NH4+ , который удаляется с мочой.
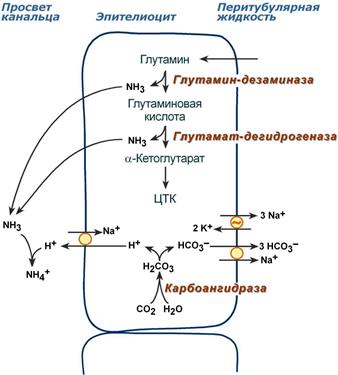
Реакции аммониегенеза
Аммониегенез происходит на протяжении всего почечного канальца, но более активно идет в дистальных отделах – дистальных канальцах и собирательных трубочках коркового и мозгового слоев. В этих сегментах секреция ионов Н+ происходит активно с участием Н+‑АТФазы, локализованной на апикальной мембране эпителиоцита.